
|
|
|
| |
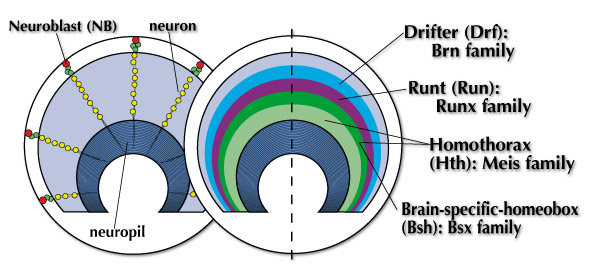
この「同心円ゾーン」は一体どのような仕組みによって形成されるのでしょうか?無数の神経細胞のうちの一部が規則正しく4種の転写因子を発現し、同心円ゾーンを形作る分子機構を明らかにするというのが第1のプロジェクトです。
|
|
 |
|
|
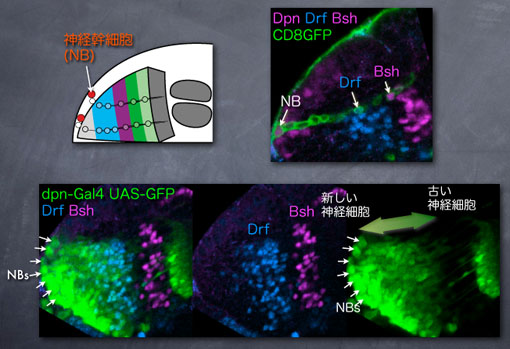
幼虫期のメダラ前駆体は半球状の構造を取っており、その表面に赤で示した神経幹細胞(neuroblast: NB)が位置しています。一つ一つの神経幹細胞は多くの神経細胞(neuron: 黄)を脳の内側に向かって直線状に生み出します。この時すでに各神経細胞がどのようなタイプのものになるかは運命づけられています。同心円ゾーンは4種の転写因子の発現によって規定されていると書きましたが、内側からMeisファミリーのHomothorax(Hth), BsxファミリーのBsh, RunxファミリーのRunt, BrnファミリーのDrfというように順に並んでいます。これら転写因子の発現は各神経細胞のタイプと一致する、つまりこれら転写因子は神経細胞タイプのマーカーであることが分かっています。
|
|
|
| |
 |
| |
 |
|
| |
同心円ゾーンが形成されるメカニズムとして、少なくとも「神経細胞の産生順」が重要な役割を果たすことが分かっています。上の図は神経細胞の産生順と転写因子の発現は相関していることを示しています。左上の図のように1つのNBから多くの神経細胞が産み出されますが、この時、1つのNBから緑・紫・青と様々なタイプの神経細胞が産み出されます。右上の写真では1つのNBから産み出される細胞がGFPでラベルされていて、1つのNBから1つのBsh陽性細胞と1つのDrf陽性細胞が産み出されていることが分かります。これら神経細胞はどのような順序でNBから産み出されたのでしょうか?
下の図ではNBから産み出された細胞がGFPでラベルされており、最近生まれた細胞ほど強くGFPが発現するようになっています。ですから、NBにより近い(画面左側)ほど新しい神経細胞が、NBからより遠い(画面右側)ほど古い神経細胞が位置しているということがわかります。そして、このGFP発現の強さとDrfやBshの発現領域が一致していることがわかります。例えばBshはGFPをほとんど発現しない古い神経細胞、Drfは強くGFPを発現する新しい神経細胞において発現しています。このことから、神経細胞の産生順と転写因子発現は相関しているということができます。
Hasegawa, E., Kitada, Y., Kaido, M., Takayama, R., Awasaki, T., Tabata, T. and Sato, M.
Concentric zones, cell migration and neuronal circuits in the Drosophila visual center.
Development (2011) 138, 983-993 (Open Acess PDF file) |
|
| 神経幹細胞は1つ1つ順に形成する |
|
|
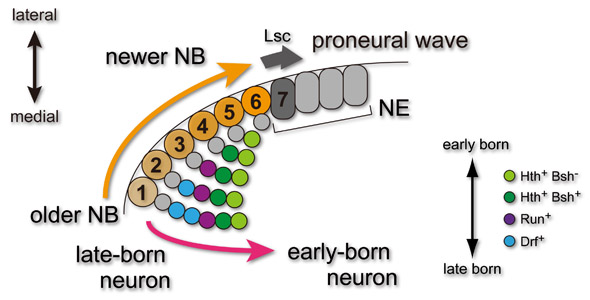
では、神経細胞の産生順と神経細胞のタイプ(転写因子の発現)はどのようにしてリンクしているのでしょうか?その前に、神経幹細胞(NB)が産み出される仕組みについて説明する必要があります。発生の初期においてメダラの表面は未分化な神経上皮(NE)で覆われています。このうち、図の左下(medial)から右上(lateral)に向かって順にNEがNBへと分化していきます。図中の①〜⑥はNBの分化した順番をあらわしています。この図では早く分化したNBほど多くのneuronを産み出していますが、最終的にはどのNBもたくさんのneuronを産むことになります。NBは時間とともに異なるタイプの神経細胞を産み出すわけですが、このようなNBの性質の変化はNBにおいて時間と共に発現変動する遺伝子によって制御されていると考えることができます。例えば、NB⑤⑥において発現する遺伝子は初期タイプの神経細胞(Hth+/Bsh+)を、NB③④において発現する遺伝子は中期タイプの神経細胞(Run+)を、NB①②において発現する遺伝子は後期タイプの神経細胞(Drf+)を産み出す働きをしていると考えられます。
|
|
|
| |
| 神経幹細胞において一過的・順番に発現する転写因子群 |
|
|
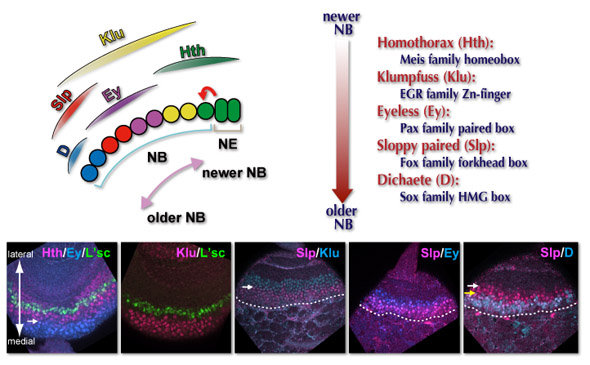
私たちは実際にNBにおいて一過的にそして順番に発現する一群の転写因子が存在することを見出しました。若いNBから順にMeisファミリーのHomothorax(Hth), EGRファミリーのKlumpfuss(Klu), PaxファミリーのEyeless(Ey), FoxファミリーのSloppy paired(Slp)そしてSoxファミリーのDichaete(D)が少しずつオーバーラップしながら発現しているのです。下の写真は実際の発現パターンの例を示しています。
|
|
|
| |
神経幹細胞において転写因子の発現変化を制御する分子機構 |
|
|
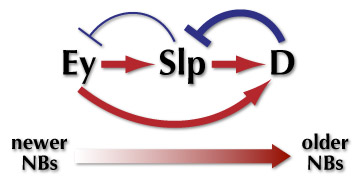
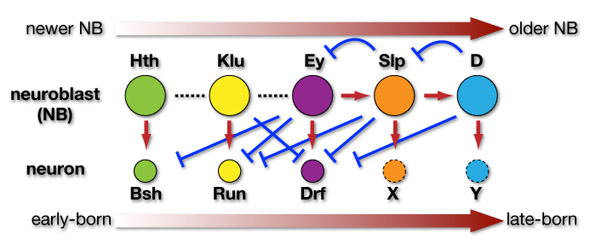
NBにおける転写因子の発現の移り変わりはどのようにして制御されているのでしょうか?少なくともEy, Dlp, Dの三者の間には上の図のような関係があることを明らかにしました。つまり、EyはSlpの発現を誘導し、SlpはEyの発現を抑えるのです。同様に、SlpはDの発現を誘導し、DはSlpの発現を抑制します。このようなことが1つのNBの中で起きると、最初にEyを発現していたNBは次第にSlp、そしてDを発現するようになると考えられます。
|
|
|
| |
| 神経幹細胞において発現する転写因子が産み出される神経細胞タイプを決める |
|
|
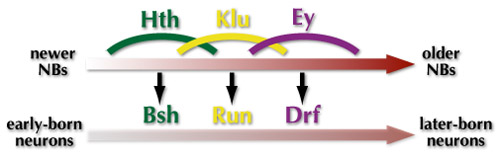
ではこれら転写因子がNBにおいて発現しているとき、各NBからはどのような神経細胞が産み出されるのでしょうか?HthとEyについてはNBだけでなく、NBから産み出される神経細胞においても持続して発現が維持されます。若いNBはHthを発現しますが、その時産み出された神経細胞はHthとBshを発現します。同様にEyを発現するNBが産み出す神経細胞はEyとDrfを発現します。Kluを発現するNBはRun陽性細胞を産み出すと考えられます。
|
|
|
| |
| 産生順依存的な神経細胞タイプ決定機構 |
|
|
まとめると、図のような相互作用によって産生順に従った神経細胞のタイプ決定が制御されていると言えます。NBにおいてHth, Klu, Ey, Slp, Dの順で転写因子が発現しますが、このうち少なくともEy, Slp, DについてはNBにおける発現の移り変わりを説明することができました。また、Hth, Klu, EyについてはそれぞれのNBにおける発現がBsh+, Run+, Drf+といったタイプの神経細胞の産生を誘導することが分かりました。実際には他にも多くの遺伝子が関与することによって神経細胞の多様性が産み出されていると考えられます。
Suzuki, T., Kaido, M., Takayama, R. and Sato, M.
A temporal mechanism that produces neuronal diversity in the Drosophila visual center.
Developmental Biology 380 (2013) 12-24.
|
|
|
| |
|