
|
|
|
| |
4種の転写因子によって規定される神経細胞は最終的にどのような神経回路を構築し、どのような機能を果たすのでしょうか?このような問題を解決するためには個々の神経細胞特異的に遺伝子発現を誘導し、その投射パターンを明らかにしたり、神経機能を人工的に操作するといった実験が必要です。その第一歩として、それぞれの遺伝子発現を正確に再現するGal4系統を作製し、GFPを用いて可視化させることによって各神経細胞の投射パターンを詳細に解析しました。
|
|
 |
|
|
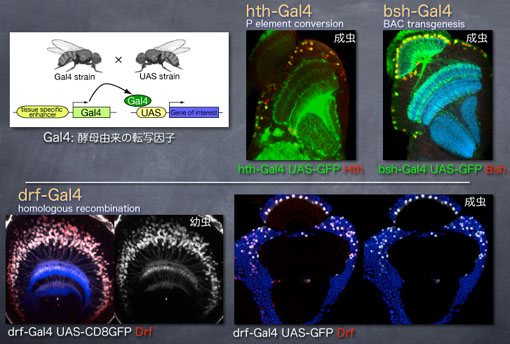
Gal4は酵母由来の転写因子で、その結合配列であるUAS配列下流に位置する任意の遺伝子発現を人工的に誘導することができます。当研究室では各転写因子の発現を正確に再現するようなGal4系統を相同組み換え、BACトランスジェニックなどの方法を駆使して作製しています。ここではHth, Bsh, Drfそれぞれの発現を再現するGal4系統としてhth-Gal4, bsh-Gal4, drf-Gal4系統の発現パターンを示しています。
|
|
|
 |
|
| |
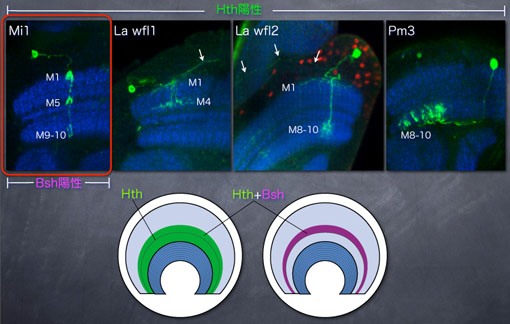
上記のGal4系統を用いた場合、多数のメダラ神経細胞がラベルされてしまい、1つ1つの神経細胞の投射パターンは判別できません。そこで、MARCMと呼ばれるモザイク解析の手法を用いて、Gal4を発現する神経細胞のうちごく一部のみをGFPでラベルしました。その結果、Hthを発現するメダラ神経細胞は形態的に4種類に分類できることがわかりました。Hth陽性細胞のうちの一部がBshを発現するのですが、興味深いことにBshは4種あるHth陽性神経細胞のうち、Mi1と呼ばれるタイプの神経細胞においてのみ特異的に発現していました。つまり、Bsh発現とMi1神経細胞は1対1対応すると言えます。
|
|
|
Bshは単一種類の神経細胞Mi1において特異的に発現している |
|
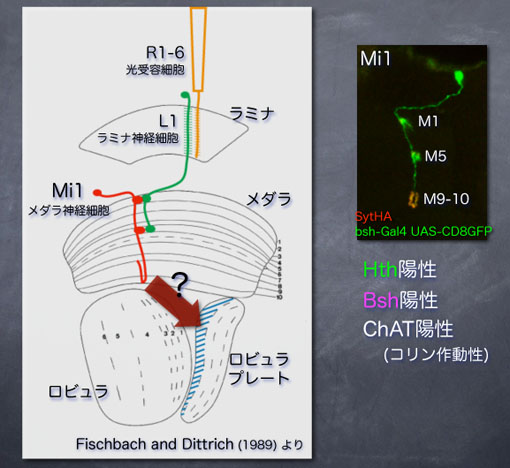
Mi1神経細胞は昔から重要な神経細胞であると言われていました。古典的な解剖学的解析から、Mi1はラミナ神経細胞であるL1からの情報を受け取ると考えられています。光受容細胞R1-6によって受け取られた光情報はL1, Mi1へと伝達されると考えられます。ここで、R1-6とL1は物体の動き、つまり動体認識に関わることが分かっていますので、Mi1は動体認識に関わる神経回路を構成していると考えることができます。
さらに、Bsh陽性細胞はマーカーの発現からコリン作動性であることが分かっているので、Bshの発現によってMi1という1種類のメダラ神経のタイプが決まり、そしてそれは蛹の時期には脳の内側から外側へ向かって移動し、コリン作動性の動体認識に関与する神経細胞になるということが言えるわけです。メダラは非常に多くの神経細胞から成りますが、たった1つの遺伝子発現によってこのような性質が一意に決まっているというのは非常に重要な知見であると言えるでしょう。
|
|
BshとHthが発現することによりMi1神経細胞の運命が決定する
|
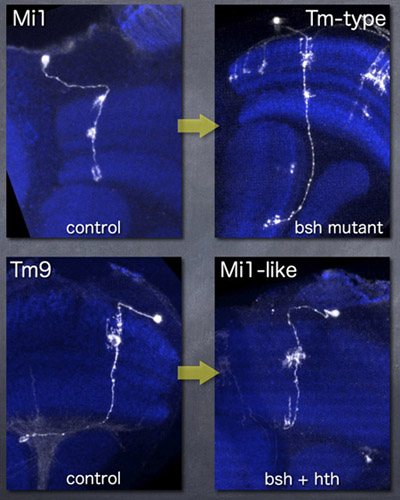
bsh遺伝子はMi1神経細胞において特異的に発現していますが、bsh変異体においてはMi1が全く別のタイプの神経細胞に変化します。コントロールにおいてMi1の投射はメダラに限局していますが、bsh変異体においてはロビュラまで投射するタイプの神経細胞になっています。一方、Mi1以外の神経細胞(例えばTm9)においてbshを強制発現すると、Mi1と似た神経細胞に変化します。Mi1はBshに加えてHthも発現しているので、bshとhthの両方を強制発現させた時の方がより本来のMi1に近い形態の神経細胞が生じます。bshとhthが協調して働くことによってMi1という神経細胞タイプが決定されると言えます。
|
|
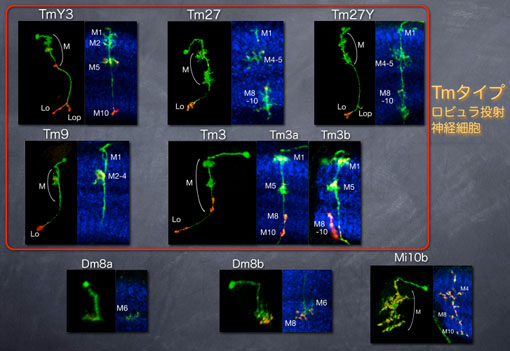
Drfは9種類のメダラ神経細胞において発現する
| |
 |
|
|
Drf陽性細胞は9種類の神経細胞に分類できます。この時、そのほとんどがメダラだけでなくロビュラに投射するTmタイプの神経細胞であることがわかりました。Hth陽性細胞は決してロビュラには投射しないので、Drf陽性細胞とHth陽性細胞は大きく異なると言うことができます。
hth, bsh, drfそれぞれの変異体を用いた解析から、hthとbshは神経細胞のタイプ決定に関わるが、drfの機能は大きく異なり、神経細胞の一部の性質、たとえば樹状突起や軸索の形成に関与することが分かりました。同心円上に発現する転写因子とは言ってもその機能は多様であり、機能分担することによって複雑な神経回路を形成していると考えることができます。
同心円ゾーンを形成する転写因子はここで述べた4つ以外にも多数存在すると考えられ、実際私たちはそのような新たな遺伝子を探索し、機能解析しています。同心円ゾーンはメダラの発生・機能を理解する上で非常に重要な基盤になると考えられます。
Hasegawa, E., Kitada, Y., Kaido, M., Takayama, R., Awasaki, T., Tabata, T. and Sato, M.
Concentric zones, cell migration and neuronal circuits in the Drosophila visual center.
Development (2011) 138, 983-993 (Open Acess PDF file)
Hasegawa, E., Kaido, M., Takayama, R. and Sato, M.
Brain-specific-homeobox is required for the specification of neuronal types in the Drosophila optic lobe.
Developmental Biology 377 (2013) 90-99.
|
|
|