
|
|
|
| |
分子生物学・分子遺伝学を用いた研究に加え、私たちの研究室では数理モデルを活用した研究にも力を入れています。これによって、通常の生命科学研究では扱うことが困難な現象について理解を深め、かつモデルから得られた予測を元に、遺伝子ネットワークの新しい機能を実験的に解明することが可能となります。
細胞と細胞が情報のやりとりをする時,長距離性の情報伝達は拡散性のタンパク質によって,短距離性の情報伝達は細胞膜上のタンパク質によって隣接する細胞に伝達されます。EGFは長距離性の,Notchは短距離性の情報伝達を担う中心的な因子として知られています。しかし,この両者が協調して働いた時にどのような効果を示すのか,通常の生命科学実験によって調べることは困難であり,ほとんど理解が進んでいませんでした。本研究では脳の形成過程において長距離性の情報伝達因子であるEGFと短距離性の情報伝達因子Notchの協調作用に注目し,数理モデリングを活用したコンピューターシミュレーションの結果を実験的に検証することによって,Notchの働きがEGF存在下では大きく変化することを見出しました。
EGFとNotchの協調作用を調べるにあたり,ショウジョウバエ脳の神経幹細胞形成過程において見られる「分化の波」Proneural Waveに注目しました。類似した分化の波は他の生物においても存在します。Notchの短距離作用が働いた場合,一般的にはいわゆるゴマシオパターンを形成しますが,Proneural Waveにおいてそのようなパターンは見られません。そこで,数理モデルに基づいたコンピューターシミュレーションを行ったところ,EGFの産生を減少させるとゴマシオパターンが現れることが予測されました。実際にEGFの産生量を減少させたところ,脳において明らかなゴマシオパターンが現れたことから,Proneural WaveにはNotchによる短距離性作用が確かに組み込まれており,EGFとの協調作用によって波の伝播速度を制御するという新たな役割を果たすことが明らかとなりました。
EGFとNotchの協調作用は大脳皮質の形成過程における神経幹細胞の分化や,肺がん・乳がんの発症過程においても重要な役割を果たしていると考えられることから,本研究によって明らかとなった遺伝子ネットワークの動作機構,およびその正確なシミュレーションを実現する数理モデルは今後これら様々な生命現象の研究に対しても応用できると期待されます。
|
|
 |
|
|
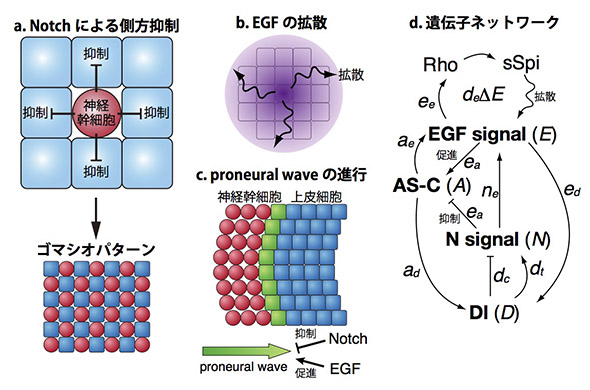
細胞と細胞が情報のやりとりをする時,長距離性の情報伝達は拡散性のタンパク質によって,短距離性の情報伝達は細胞膜上のタンパク質によって隣接する細胞に伝達されます。EGFは長距離性の,Notchは短距離性の情報伝達を担う中心的な因子として知られています。Notchの短距離作用は側方抑制と呼ばれ,例えば図のaにおいて青い細胞が赤い細胞に変化(分化)する時,赤い細胞は隣り合う細胞に対して抑制性のシグナルを送り,それによって赤い細胞の隣りには必ず青い細胞が存在するようなゴマシオパターンを形成します。一方EGFは拡散性因子であり,産生された領域から離れた場所において長距離作用を及ぼします(図のb)。NotchとEGFはどちらも様々な生命現象において重要な役割を果たしますが,Notchによる側方抑制とEGFの拡散効果が協調して働いた時にどのような効果をもたらすのか,明らかにされていませんでした。
本研究ではこのような協調作用の働きを研究するためにショウジョウバエの脳において見られるProneural Waveという現象に着目しました。形成過程のハエの脳においてはまず青い上皮細胞のみが存在していますが,これが1列ずつ順番に赤い神経幹細胞に変化します (図のc)。このような「分化の波」Proneural Waveの進行はEGFによって促進され,Notchによって抑制されることが知られていました。
Notchによる側方抑制はあらゆる動物において見られる現象であり,Proneural WaveにおいてもNotchが側方抑制を引き起こすための条件は揃っています。Notchによる側方抑制が働けば図のaのようなゴマシオパターンが生じるはずですが,実際にはProneural Waveにおいてはそのようなパターンは見られません(図のc)。果たしてNotchは側方抑制を引き起こしているのでしょうか?もしそうだとすれば,なぜNotchはゴマシオパターンを形成せずに波の進行を制御できるのでしょうか?
通常の生命科学実験のみではこのような疑問に答えることはできません。そこで,本研究ではNotchとEGFによるProneural Waveの制御についての数理モデルを構築し,コンピューターシミュレーションと生命科学実験を組み合わせることによってNotchとEGFによる遺伝子ネットワークの働きを明らかにしました(図のd)。
|
|
|
 |
|
| |
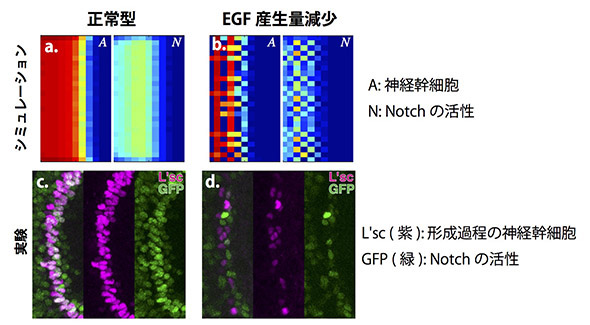
我々はまずProneural WaveがNotchによる側方抑制を含んでいると仮定し,EGFの拡散とNotchによる側方抑制を組み合わせたProneural Waveの数理モデルを構築しました。興味深いことに,この数理モデルは生体内で見られるProneural Waveと非常に近い挙動を示しました(図a, c)。つまり,通常はゴマシオパターンを示さずに分化の波が進行する上,EGFの活性をなくしたEGF変異体においては波が消失し,Notchの活性をなくしたNotch変異体では波の進行が加速するなど,様々な変異体の状況も的確に再現したのです。EGFの拡散とNotchの側方抑制だけでProneural Waveの挙動を全く矛盾無く説明できたと言えます。
では,ゴマシオパターンは一体どこに行ってしまったのでしょうか?EGFは拡散性因子であり,Notchがゴマシオ状に活性化していたとしても,EGFの拡散効果によってそれが隠されてしまう可能性が考えられました。そこでシミュレーション上でEGFの拡散もしくは産生量を減少させたところ,そのような状況下ではゴマシオパターンが現れることが予測されました(図b)。EGFの活性が完全に無くなってしまうとそもそも波が消失してしまうため,我々はRNAiと呼ばれる技術によってEGFの活性を中程度に減少させることを試みました。その結果,Notchの活性および神経幹細胞の形成パターンがゴマシオ状に変化し,生体内においてもシミュレーション上と同じような互い違いのパターンが観察されました(図d)。これらの結果から我々の数理モデルが正しいこと,そしてNotchによる側方抑制が確かに生体内においてProneural Waveに含まれていることが示されました。
|
|
|

| |
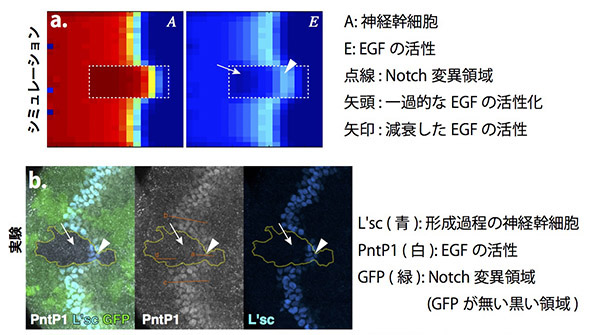
私たちはこの数理モデルを用いてさらにもう一つの問題を解決しようと試みました。Notchの活性が失われたNotch変異領域では波の進行が加速しますが,この時EGFの活性が失われることが知られていました。EGFは波の進行において必須な因子であり,EGFが失われると波も失われるはずです。Notch変異領域ではEGFが失われているにもかかわらずなぜ波が消失せずにむしろ加速するのか,これまでの研究において解決されていない疑問点でした。
私たちは上記の数理モデルにおいてNotch変異領域を設定し(図a点線内),どのようなことが起きているか観察しました。興味深いことに,Notch変異領域において波が加速する際,一過的にEGFが活性上昇し,その後減衰することが分かりました。Notch自体は神経幹細胞の形成を阻害するので,EGFが活性化し,かつNotchが失われた状況では神経幹細胞の形成は加速するはずであり,これがNotch変異領域において波の進行が加速する原因であると考えられました。これと同じことが生体内でも起きているか調べたところ,シミュレーションと同様,EGFの活性が一過的に上昇していることが確認されました(図b)。このように,数理モデルを用いることによって従来の生命科学実験だけでは解決できない問題が解決されたのです。
|
|
|
Jak/Statは脳の形成過程において生じるノイズの効果を除去し、
正確な神経幹細胞の分化を保証する
| |
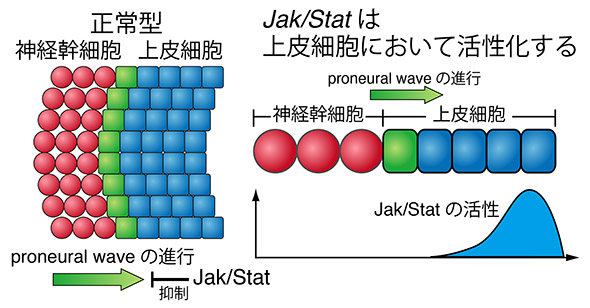
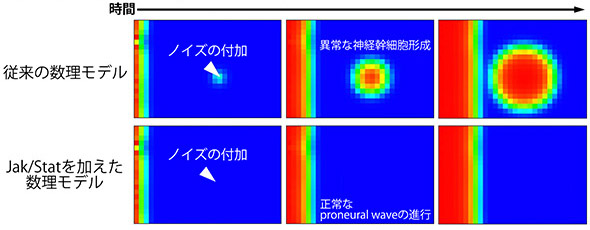
EGFとNotch以外にもJak/Statと呼ばれる細胞内情報伝達経路がProneural Waveの進行を抑制することが遺伝学的な解析からすでに報告されていました(Yasugi et al., Development, 135: 1471-80, 2008.)。Jak/Statは未分化な上皮細胞において活性化し、この働きが無くなると波の進行が加速します。ですが、数理モデルと遺伝学実験を組み合わせた解析によって、このJak/Statが脳の形成過程において乗じるノイズの効果を除去するというさらなる役割を持っていることを明らかにしました。
|
|
ノイズに弱い数理モデルにJak/Statを加えると、
ノイズに強くなった
| |
従来の数理モデルは ノイズに弱いという欠点がありました。上図のように、未分化な上皮細胞の領域に弱いノイズを加えるだけで神経幹細胞が爆発的に分化してしまうのです。ですが、実験事実に基づいてJak/Statの働きを数理モデルに加えると、ノイズによる効果が弱められ、正常なproneural waveの進行が再び見られるようになったのです(下図)。このことはJak/Statがノイズをキャンセルする働きを持っていることを示唆します。このようなJak/Statの働きを検証するため、以下のような実験を行いました。
|
|
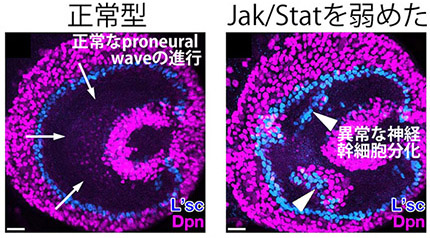
生体内においてJak/Statの働きを弱めると、
神経幹細胞が乱雑に分化した
| |
通常は、左図のようにProneural Waveは脳の外側から内側に向かって均一に進行します。分化しつつある水色でラベルされた細胞はきれいに円周上に並んでいることがわかります。ですが、上皮細胞においてJak/Statの働きを均一に弱めると、それによって神経幹細胞が乱雑に分化し、均一なproneural waveのパターンが見られなくなってしまいました。このように、数理モデルを用いた解析と実際の脳を用いた実験から、Jak/Statが脳の形成過程においてノイズの影響を除去する働きを持っていることが明らかになりました。
|
|
生命科学と数理科学の融合研究は世界的な潮流となっていますが,生命科学実験とシミュレーションを1対1に対応させて研究を推進する真の異分野融合研究はいまだ困難です。本研究では実際の生命現象に合致した数理モデルを構築し,その正しさを実験によって検証しました。
EGFとNotchの協調作用は哺乳類大脳皮質の形成過程における神経幹細胞の分化,肺がん・乳がんの発症過程においても重要な役割を果たしていると考えられています。また、Jak/Statも進化的に保存されたシグナル伝達系で,ショウジョウバエだけではなくマウスやヒトの幹細胞においても重要な役割を果たします。従って,哺乳類の大脳皮質における神経幹細胞や,万能幹細胞であるES細胞,および他のさまざまな臓器の幹細胞においても同様にノイズを除去している可能性が考えられます。本研究によって明らかとなったEGF, Notch, Jak/Statの作用機構およびその正確なシミュレーションを実現する数理モデルはヒトの脳の形成機構の解明,がんの進行をコントロールする技術の創出など,従来の生命科学研究だけでは解決できないような複雑な問題の解明に役立つと期待されます
Sato, M., Yasugi, T., Minami, Y., Miura, T. and Nagayama, M.
Notch-mediated lateral inhibition regulates proneural wave propagation when combined with EGF-mediated reaction diffusion. Proceedings of the National Academy of Sciences 113, E5153-E5162 (2016).
Tanaka, Y., Yasugi, T.,Nagayama, M., Sato, M. and Shin-Ichiro Ei.
JAK/STAT guarantees robust neural stem cell di erentiation by shutting off biological noise.
Scientific Reports 8, 12484 (2018).
|